Inquiry Basket
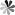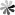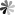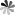 Loading......
Loading......
Research on cancer treatment has made significant progress in the past few decades, and new anti-cancer drugs are constantly emerging. Microtubule inhibition is now one of the most important mechanisms of cytotoxic drugs in the current generation. Although tubulin stabilizers (e.g., Paclitaxel) and destabilizers (e.g., Vincine) have been clinically successful as stand-alone anticancer agents, they may cause systemic toxicity in many patients. To minimize the systemic toxicity, scientists combined tubulin inhibitors with antibodies to target tumors, a concept that eventually evolved into antibody-conjugated drugs known as "magic bullets".
Microtubules are hollow filaments composed of alpha-tubulin and beta-tubulin. Like actin polymers and intermediate fibers, microtubules are components of the cytoskeleton and are essential for a variety of cellular functions, including maintenance of cell shape, cell movement, and intracellular transport of vesicles, mitochondria, and other components. Microtubules play an important role in cell division in the context of cell proliferation. During cell division, the entire microtubule network rearranges to form the mitotic spindle, thus providing the structural framework for the physical segregation of sister chromatids. The dynamic nature of microtubules is essential for the optimal arrangement of mitotic spindles, as well as for the subsequent process of sister chromatid movement. Thus, drugs that interfere with microtubule dynamics can have significant effects on spindle formation and the eventual cell division process. Currently, the most important tubulin inhibitors used as ADC payloads are Auristatin, Eribulin, which are derived from marine species, and plant-derived Maytansinoids.
 Fig. 1. Microscopic image of a cell during mitosis, in which green tubulin helps separate the red chromosomes.
Fig. 1. Microscopic image of a cell during mitosis, in which green tubulin helps separate the red chromosomes.
The parent structure of Auristatin is Dolastatin 10, a toxin isolated from Dolabella auricularia. The synthetic Dolastatin 10 analogs monomethyl auristatin E (MMAE) and monomethyl auristatin F (MMAF) are widely used as payload components of ADCs. Brentuximab vedotin (Adcetris®), for example, binds MMAE to a CD30-targeted antibody with partially reduced interchain disulfide bonds via a maleimide-valine-citrulline linker. CD30 is a cell membrane antigen expressed by tumor cells of patients with systemic anaplastic large cell lymphoma (sALCL) and classical Hodgkin's lymphoma (HL). Originally approved by the FDA in 2011, Adcetris® has become the first new treatment for Hodgkin's lymphoma since 1977 and the first treatment specifically targeting sALCL.
 Fig. 2. Structure of Brentuximab vedotin (Adcetris®).
Fig. 2. Structure of Brentuximab vedotin (Adcetris®).
Maytansinoids are a class of benzoansamacrolides, which were first extracted from Maytenus boaria. It can bind to tubulin at the Rhizoxin binding site to inhibit the formation of microtubules. New derivatives, such as DM1, DM3, and DM4, are widely used in clinical ADC structures. These derivatives differ only in the number of methyl groups near the sulfhydryl binding site. Among currently marketed ADCs, Trastuzumab emtansine (Kadcyla®) combines the anti-HER2 monoclonal antibody Trastuzumab (Herceptin®) with DM1 via a succinimidyl 4-(N-maleimidomethyl)cyclohexane-1-carboxylate (SMCC) linker. Kadcyla is also the first ADC approved for use in solid tumors.
 Fig. 3. Structure of Trastuzumab emtansine (Kadcyla®).
Fig. 3. Structure of Trastuzumab emtansine (Kadcyla®).
The most widely used tubulin inhibitors are Auristatin and Maytansinoids while other tubulin inhibitors have also been applied in ADCs, including Taxol, Vincristine, and Colchicine derivatives. Recently, Eribulin is added as a toxin molecule in the ADC drug MORAb-202 and has entered the clinical stage.
Currently, many other molecules with different structures that inhibit tubulin have been found to have the potential to act as ADC payloads. Studies have shown that Dictyostatin can inhibit the growth of certain tumor cells. Several analogs of Dictyostatin have been designed and synthesized, and they show significant cytotoxicity in several taxane-resistant tumor cell lines. However, it is currently unclear whether Dictyostatin and Paclitaxel have similar binding sites on tubulin.
 Fig. 4. Structures of Dictyostatin, Indibulin, Cevipabulin, and Cryptophycin-52.
Fig. 4. Structures of Dictyostatin, Indibulin, Cevipabulin, and Cryptophycin-52.
 Fig. 5. Structures of Rhizoxin, and Tubulysin A.
Fig. 5. Structures of Rhizoxin, and Tubulysin A.
Although ADC payloads are dominated by Auristatin and Maytansine compounds with tubulin targeting mechanisms, other tubulin inhibitors have been evaluated such as vinca alkaloids and taxanes. To date, none of these drugs have made it to the clinical stage. Only EISAI recently used Eribulin as a payload for MORAb-202, an ADC currently being evaluated in the clinical phase. The favorable clinical results of MORAb-202 may enable further development of ADCs in the future.










 Loading......
Loading......